Дмитрий Азаров - В поисках прародины индоевропейцев. Теория сдвигов согласных звуков
Здесь есть возможность читать онлайн «Дмитрий Азаров - В поисках прародины индоевропейцев. Теория сдвигов согласных звуков» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. ISBN: , Издательство: Литагент Ридеро, Жанр: Языкознание, История, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.
- Название:В поисках прародины индоевропейцев. Теория сдвигов согласных звуков
- Автор:
- Издательство:Литагент Ридеро
- Жанр:
- Год:неизвестен
- ISBN:9785447491024
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
В поисках прародины индоевропейцев. Теория сдвигов согласных звуков: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «В поисках прародины индоевропейцев. Теория сдвигов согласных звуков»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
В поисках прародины индоевропейцев. Теория сдвигов согласных звуков — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «В поисках прародины индоевропейцев. Теория сдвигов согласных звуков», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
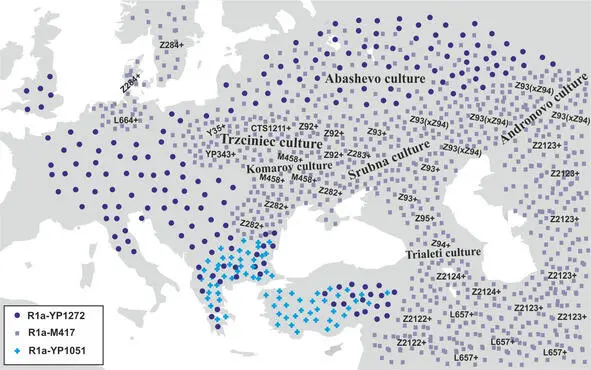
Рис.16 Географическая локализация носителей субкладов R1a-YP1272 , R1a-YP1051 и R1a-M417 в период с 2200 по 1600 гг. до н.э.
Примерно в 1600 году до н.э. носители субклада R1a-YP343 мигрировали с территории тшинецкой культуры далее на запад и вторглись на территорию унетицкой культуры. В результате этого вторжения унетицкая культура сменяется культурой курганных погребений. Носители субкладов R1a-Z282* мигрируют несколькими волнами с территории сегодняшней Румынии на юг Балканского полуострова и на территорию Анатолии. Примерно в это же время на территории Греции возникает микенская цивилизация. Экспансия носителей субкладов R1a-Z282* приводит к миграции населения Балканского полуострова на территорию Анатолии и Ближнего Востока (миграция «народов моря»). Часть населения срубной культуры (носителей субкладов R1a-Z93* ) смещаются на запад на территорию Украины и Молдавии. Географическая локализация основных субкладов R1a к концу II-го тысячелетия до н.э. была примерно следующей (рис.17).
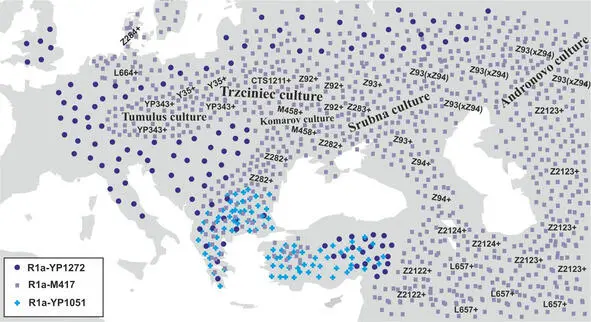
Рис.17 Географическая локализация носителей субкладов R1a-YP1272 , R1a-YP1051 и R1a-M417 в конце II-го тыс. до н.э.
Основной этап расселения носителей y-гаплогруппы R1a закончился к концу II-го тыс. до н. э. Дальнейшие миграции носителей y-гаплогруппы R1a не были столь существенными и не привели к заметным изменениям структуры населения на территории Евразии.
Таким образом, в упрощенном виде миграции носителей y-гаплогруппы R1a можно схематично представить как последовательный выход трех крупных групп носителей данной y-гаплогруппы с территории Иранского нагорья, их разделение на территории Ближнего Востока в период раннего неолита, проникновение на территорию Европы и миграции в Центральную и Западную Европу в период бронзового и железного веков (рис.18).
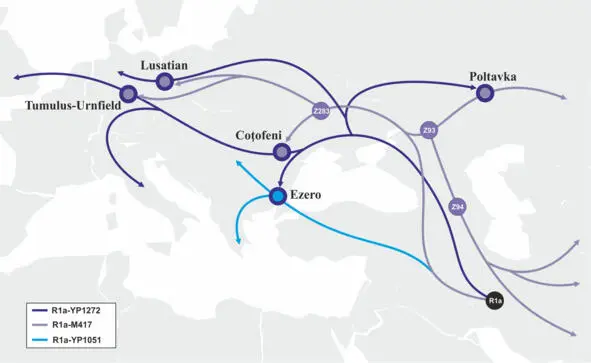
Рис.18 Схема миграций носителей y-гаплогруппы R1a
Глава II. Теория сдвигов согласных звуков
2.1. Основная идея теории сдвигов согласных звуков
Использование информации о расселении носителей определенных y-гаплогрупп может существенно упростить моделирование процессов генезиса протоязыков. ДНК генеалогия дает возможность с достаточно высокой точностью определить популяции, которые могли участвовать в генезисе определенных протоязыков, а также установить временные и географические границы этих процессов. Информация о популяциях, принимавших участие в генезисе протоязыков, дает возможность установить связь между исходными языками популяций, принимавших участие в генезисе, и языками, возникшим в результате смешения этих популяций. Другими словами, если мы знаем, что язык AB является результатом смешения популяций A и B, мы можем сделать определенные предположения относительно исходных языков, на которых говорили популяции A и B до их смешения (рис.19).
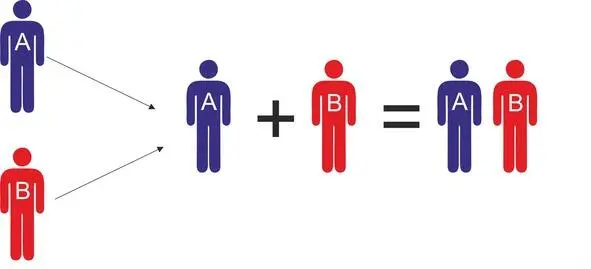
Рис. 19 Схема смешения популяций
Если мы посмотрим на филогеническое дерево y-гаплогрупп, то увидим, что разделение на основные y-гаплогруппы произошло примерно ~65 000 – ~47000 лет назад. Информация о географической локализации носителей различных y-гаплогрупп дает возможность оценить вероятность контактов между носителями определенных y-гаплогрупп или об отсутствии таковых. Примерный маршрут миграции АСЧ после выхода с территории Африки может быть восстановлен на основании данных о распределении субкладов y-гаплогрупп в современных популяциях (рис.20).
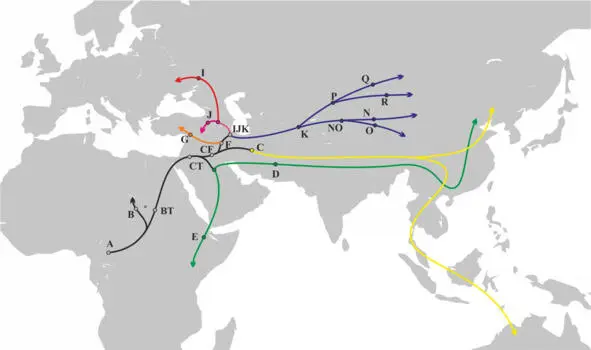
Рис.20 Маршруты миграции АСЧ после выхода с территории Африки
Определив носителей y-гаплогрупп, которые могли иметь между собой контакты после разделения на соответствующие y-гаплогруппы, можно определить, какие лингвистические макроветви должны были возникнуть после первичного расселения АСЧ. С учетом географической локализации y-гаплогрупп и времени их разделения на отдельные y-гаплогруппы, все вновь возникшие в ходе миграций АСЧ в период с ~70 000 по ~45 000 гг. до н.э. языки можно разделить на пять макроветвей (рис.21).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «В поисках прародины индоевропейцев. Теория сдвигов согласных звуков»
Представляем Вашему вниманию похожие книги на «В поисках прародины индоевропейцев. Теория сдвигов согласных звуков» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «В поисках прародины индоевропейцев. Теория сдвигов согласных звуков» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.