Инал Акоев - Биофизика познает рак
Здесь есть возможность читать онлайн «Инал Акоев - Биофизика познает рак» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 1988, Издательство: Наука, Жанр: sci_biophys, Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Биофизика познает рак
- Автор:
- Издательство:Наука
- Жанр:
- Год:1988
- Город:Москва
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Биофизика познает рак: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Биофизика познает рак»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Книга предназначена для широкого круга читателей, интересующихся современными вопросами медицины и биологии.
Биофизика познает рак — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Биофизика познает рак», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Таким образом, обобщая данные о состоянии кроветворения в период длительного слабого хронического воздействия радиации и в период отдаленного пострадиационного восстановления после острых и многократных облучений, можно заключить, что наиболее характерным для гемопоэза было явное или скрытое напряжение эритропоэза, в значительной мере определявшее состояние периферической крови и клеточность костного мозга. Значительную роль при этом играет сокращенная продолжительность жизни эритроцитов. Основной вклад в пополнение клеточности костного мозга после экстремальных воздействий вносят процессы пролиферации и дифференцировки морфологически не различимых стволовых клеток костного мозга. Они обладают чрезвычайными потенциями к пролиферации и имеют другие возможности усиления продуктивности отдельных ростков кроветворения.
Регуляция размножения и дифференцировки кроветворных клеток
В связи с успехами исследования процессов кроветворения и функциональных возможностей стволовых клеток при использовании метода селезеночных колоний и культур на полутвердых средах и других методов представления о регуляции путей их пролиферации и дифференцировки существенно изменились.
Экспериментально подтвердилось существование единой полипотентной стволовой клетки, способной к дифференцировкам по всем направлениям гемопоэза, включая лимфопоэз (рис. 3). Казалось бы, это непосредственно доказывает правильность исходного положения о конкуренции различных ростков кроветворения за направление дифференцировки стволовой клетки в соответствии с потребностями организма. Однако вопрос оказался значительно сложнее.
Наряду с исходной полипотентной стволовой кроветворной клеткой могут существовать и обладающие стволовыми свойствами предшественники, коммитированные (т. е. ограниченные к выбору направления дифференцировки) предшественники двух типов — миелопоэза и лимфопоэза. Это первое и главное разделение направлений возможной дифференцировки единой полипотентной стволовой клетки. Как осуществляется регуляция выбора дифференцировки — в сторону миелопоэза или лимфопоэза, не очень ясно.
Стволовые клетки, коммитированные в сторону лимфопоэза, дифференцируются сначала в общих, а затем в раздельных предшественников Т- и В-лимфоцитов, из которых через ряд промежуточных стадий образуются Т-лимфоциты (приобретая определенные клеточные рецепторы) и В-лимфоциты (приобретая рецепторы к комплементу, к фрагментам и комплексам иммуноглобулинов и комплексам антиген—антитело). Кроме того, из них образуются плазматические клетки. Лимфопоэз наименее изучен. До сих пор неясно, имеются ли в лимфоцитарном ряду клетки, не способные к пролиферации. Среди лимфоидных клеток трудно определить принадлежность их к пролиферирующему, созревающему или функциональному пулам, так как возможны взаимные перекрытия.
Стволовые клетки, коммитированные в сторону миелопоэза, в дальнейшем через поэтинчувствительные предшественники разделяются на три ростка: эритроцитарный, гранулоцито-макрофагальный и тромбоцитарный.
В эритроцитарном направлении выявлено несколько промежуточных клеток (в том числе и бурстообразующая клетка) между общим предшественником миелопоэза и эритропоэтинчувствительными клетками. Последующие морфологически различаемые клетки относятся к проэритробласту, трем видам эритробластов, ретикулоциту и зрелому эритроциту. Обычно происходит 5—6 митозов в морфологически различимых клетках, и каждый эритробласт дает 30—60 эритроцитов. Эритробласты теряют способность к делению, как только в клетке накопится 27 нг гемоглобина. В случае замедления синтеза гемоглобина увеличивается число делений эритробластов. Из одной полипотентной стволовой клетки, начавшей дифференцировку в сторону миелопоэза, может образоваться около 1 млн эритропоэтинчувствительных клеток и около 0,1 млн клеток — предшественников гранулоцитов и макрофагов.
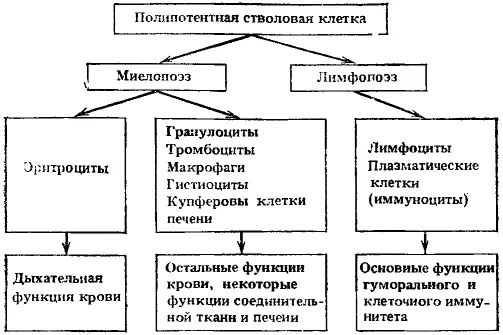
Рис. 3. Основные пути дифференцировки полипотентной стволовой клетки, клеточные формы и их главные функции
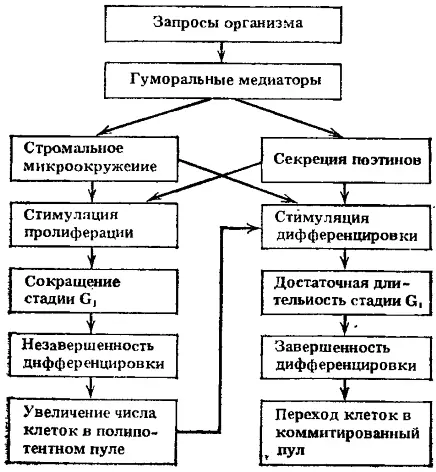
Рис. 4. Возможные пути регуляции пролиферации и дифференцировки полипотентных стволовых клеток в соответствии с запросами организма
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Биофизика познает рак»
Представляем Вашему вниманию похожие книги на «Биофизика познает рак» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Биофизика познает рак» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.