Мирослав Одинак - Клиническая диагностика в неврологии
Здесь есть возможность читать онлайн «Мирослав Одинак - Клиническая диагностика в неврологии» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. ISBN: , Жанр: Медицина, Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Клиническая диагностика в неврологии
- Автор:
- Жанр:
- Год:неизвестен
- ISBN:978-5-299-00417-5
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Клиническая диагностика в неврологии: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Клиническая диагностика в неврологии»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Руководство предназначено для врачей-неврологов, нейрохирургов, терапевтов, психиатров.
Клиническая диагностика в неврологии — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Клиническая диагностика в неврологии», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Центральные отростки первых нейронов следуют в составе заднего чувствительного корешка в задний рог спинного мозга, где они заканчиваются на дендритах клеток второго нейрона, расположенных в собственном ядре заднего рога. Большинство аксонов второго нейрона переходят через переднюю белую спайку на противоположную сторону и далее вступают в боковой канатик, составляя здесь латеральный спиноталамический путь (tractus spinothalamicus lateralis, или lemniscus spinalis) .
Данный перекрест имеет две анатомические особенности:
– переход перекрещивающихся волокон происходит в передней белой спайке не в горизонтальной плоскости, а косо вверх. В результате аксоны второго нейрона входят в латеральный спиноталамический путь противоположной стороны на 2 – 3 сегмента выше их выхода из задних рогов спинного мозга;
– перекрест происходит таким образом, что волокна от каждого последующего (нижерасположенного) сегмента вступают в латеральный спиноталамический путь снаружи. В результате наружную часть этого пути составляют волокна, идущие от нижележащих сегментов, а внутреннюю часть – от вышележащих сегментов (закон эксцентрического расположения более длинных путей Ауэрбаха – Флатау).
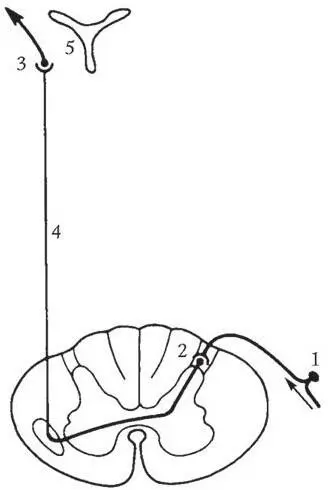
Рис. 1.1. Схема хода волокон поверхностной чувствительности (боль, температура, частично тактильные ощущения):
1 – первый нейрон; 2 – второй нейрон; 3 – третий нейрон; 4 – латеральный спиноталамический путь; 5 – таламус
После прохождения в боковых канатиках спинного мозга латеральный спиноталамический путь вступает в головной мозг, где он транзитом проходит практически через весь ствол и заканчивается лишь на клетках латеральных ядер зрительного бугра – телах третьих нейронов.
Аксоны третьего нейрона проходят через заднее бедро внутренней капсулы и далее образуют лучистый венец. Здесь волокна пути веерообразно расходятся, достигая коры головного мозга – задней центральной извилины (первичное корковое поле чувствительного анализатора) и прилежащих к ней передних отделов теменных долей.
Вышеописанная схема трехнейронного пути поверхностной чувствительности является классической. Между тем немаловажную роль играет и другой путь. Он образован частью аксонов вторых нейронов, которые вступают в передние канатики своей и противоположной стороны. В итоге формируются передний (центральный) спиноталамический и спиноретикулоталамический тракты. После восхождения по спинному мозгу в пределах продолговатого мозга они присоединяются к другим чувствительным проводникам. Передний спиноталамический путь несет импульсы к центральным и парацентральным ядрам зрительного бугра, а спиноретикулоталамический – к ретикулярным ядрам таламуса после многочисленных переключений в ретикулярной формации. От центрального и парацентрального ядер аксоны третьего нейрона направляются в поясную извилину. Данные пути несут лишь качественную информацию, определяющую модальность стимулов без тонкой характеристики и четкой локализации. С другой стороны, тонкая характеристика и четкая локализация стимула являются прерогативой филогенетически более молодого латерального спиноталамического пути. В отличие от классической схемы многие вторые нейроны этих путей не доходят до зрительного бугра, а заканчиваются на клетках центрального серого вещества среднего мозга. Отсюда по нисходящим путям импульс идет к клеткам передних рогов, клеткам ретикулярной формации ствола, клеткам коры мозжечка.
1.3. Пути проведения глубокой чувствительности
Информация о глубокой чувствительности воспринимается проприоцепторами, а расположение тел первых нейронов и их периферических отростков аналогично пути поверхностной чувствительности (рис. 1.2).
Центральные отростки рецепторных нейронов также следуют в спинной мозг через задние корешки, но, в отличие от пути поверхностной чувствительности, не заходят в серое вещество спинного мозга, а образуют задние канатики своей стороны. Здесь аксоны первого нейрона делятся на короткие и длинные ветви. Короткие ветви замыкаются на уровне своего, а также ближайших выше- и нижележащих сегментов, образуя тем самым большую часть афферентного входа для глубоких рефлексов. Из коротких ветвей также формируются коллатерали к заднему и переднему спиномозжечковым путям – задний спиномозжечковый путь (Флексига) и передний спиномозжечковый путь (Говерса), которые обеспечивают поступление части афферентной импульсации от проприоцепторов в мозжечок с последующим синергичным (согласованным) и экономным сокращением мышц-синергистов, а также расслаблением мышц-антагонистов.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Клиническая диагностика в неврологии»
Представляем Вашему вниманию похожие книги на «Клиническая диагностика в неврологии» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Клиническая диагностика в неврологии» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.