Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных
Здесь есть возможность читать онлайн «Коллектив авторов - Общая вирусология с основами таксономии вирусов позвоночных» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Год выпуска: 2012, Издательство: Литагент БИБКОМ, Жанр: Медицина, Детская образовательная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Общая вирусология с основами таксономии вирусов позвоночных
- Автор:
- Издательство:Литагент БИБКОМ
- Жанр:
- Год:2012
- ISBN:нет данных
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Общая вирусология с основами таксономии вирусов позвоночных: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Общая вирусология с основами таксономии вирусов позвоночных»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Общая вирусология с основами таксономии вирусов позвоночных — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Общая вирусология с основами таксономии вирусов позвоночных», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Синтез и внутриклеточный транспорт гликопротеидов характеризуется рядом особенностей, присущих клеточным внутримембранным белкам. Их синтез осуществляется на полисомах, ассоциированных с мембранами, и белки сразу же после синтеза попадают в шероховатые мембраны, откуда транспортируются в мембраны эндоплазматической сети и в комплекс Гольджи, где происходит модификация и комплектование углеводной цепочки, а затем – в плазматическую мембрану в ряде случаев путем слияния с ней везикул комплекса Гольджи. Такой целенаправленный транспорт осуществляется благодаря имеющейся на аминоконце белка специфической последовательности от 20 до 30 аминокислот (сигнальному пептиду). Сигнальный пептид отрезается от белковой молекулы после того, как гликопротеид достигает плазматической мембраны.
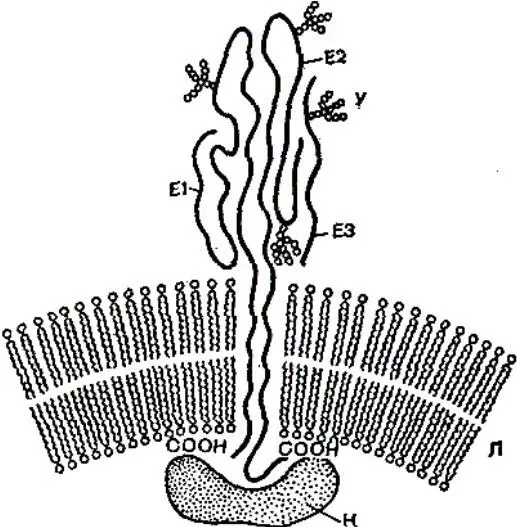
El, E2, ЕЗ – молекулы вирусных гликопротеидов; К – капсидный белок; У – углеводные цепочки; Л – липидный бислой.
Рисунок 24 – Строение липопротеидной оболочки вируса Синдбяс
Гликозилирование полипептидов является сложным многоступенчатым процессом, первые этапы которого начинаются уже в процессе синтеза полипептидов, и первый сахар присоединяется к полипептидной цепи, еще не сошедшей с рибосомы. Последующие этапы гликозилирования происходят путем последовательного присоединения сахаров в виде блоков к углеводной цепочке в процессе транспорта полипептида к плазматической мембране. Окончательное формирование углеводной цепочки может завершаться на плазматической мембране перед сборкой вирусной частицы. Процесс гликозилирования не влияет на транспорт полипептида к плазматической мембране, но имеет существенное значение для экспрессии биологической активности белка. При подавлении гликозилирования соответствующими ингибиторами (аналоги Сахаров типа 2-дезоксиглюкозы, антибиотик туникамицин) нарушается синтез полипептидов, блокируется сборка вирионов миксовирусов, рабдовирусов, альфавирусов или образуются неинфекционные вирионы герпеса и онковирусов.
Сульфирование. Некоторые белки сложно устроенных РНК- и ДНК-содержащих вирусов сульфируются после трансляции. Чаще всего сульфированию подвергаются гликопротеиды, при этом сульфатная группа связывается с сахарным компонентом гликопротеида.
Ацилировалие. Ряд гликопротеидов сложно устроенных РНК-содержащих вирусов (НА2 вируса гриппа, белок G вируса везикулярного стоматита, белок HN вируса ньюкаслской болезни и др.) содержат ковалентно связанные 1-2 молекулы жирных кислот.
Нарезание. Многие вирусные белки и в первую очередь гликопротеиды приобретают функциональную активность лишь после того, как произойдет их нарезание в специфических точках протеолитическими ферментами. Нарезание происходит либо с образованием двух функциональных белковых субъединиц (например, большая и малая субъединицы гемагглютинина вируса гриппа, два гликопротеида, Е 2и Е 3, вируса леса Семлики) либо с образованием одного функционально активного белка и неактивного фрагмента, например, белки F и HN парамиксовирусов. Нарезание обычно осуществляется клеточными ферментами. У многих сложно устроенных вирусов животных, имеющих гликопротеид, нарезание необходимо для формирования активных прикрепительных белков и белков слияния и, следовательно, для приобретения вирусом способности инфицировать клетку. Лишь после нарезания этих белков вирусная частица приобретает инфекционную активность. Таким образом, можно говорить о протеолитической активации ряда вирусов, осуществляемой с помощью клеточных ферментов.
Фосфорилирование. Фосфорпротеиды содержатся практически в составе всех вирусов животных, РНК- и ДНК-содержащих, просто и сложно устроенных. В составе большинства вирусов обнаружены протеинкиназы, однако фосфорилирование может осуществляться как вирусными, так и клеточными ферментами. Обычно фосфорилируются белки, связанные с вирусным геномом и осуществляющие регулирующую роль в его экспрессии. Одним из примеров является фосфорирование белка онкогенных вирусов, обусловливающего клеточную трансформацию. Этот белок является продуктом гена Src и одновременно протеинкиназой и фосфопротеидом, т.е. способен к самофосфорилированию.
С процессом фосфорилирования связан механизм антивирусного действия интерферона. В зараженных вирусом клетках интерферон индуцирует синтез протеинкиназы, которая фосфорилирует субъединицу инициирующего фактора трансляции ЭИФ-2, в результате чего блокируется трансляция вирусных информационных РНК. Фосфорилирование белков играет регулирующую роль в транскрипции и трансляции вирусных иРНК, специфическом узнавании вирусных иРНК рибосомой, белокнуклеиновом и белок-белковом узнавании на стадии сборки вирусных частиц.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Общая вирусология с основами таксономии вирусов позвоночных»
Представляем Вашему вниманию похожие книги на «Общая вирусология с основами таксономии вирусов позвоночных» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Общая вирусология с основами таксономии вирусов позвоночных» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.