Леонид Вишняцкий - Неандертальцы - история несостоявшегося человечества
Здесь есть возможность читать онлайн «Леонид Вишняцкий - Неандертальцы - история несостоявшегося человечества» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Санкт Петербург, Год выпуска: 2010, ISBN: 2010, Издательство: Нестор-История, Жанр: История, Биология, Прочая научная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Неандертальцы: история несостоявшегося человечества
- Автор:
- Издательство:Нестор-История
- Жанр:
- Год:2010
- Город:Санкт Петербург
- ISBN:978-5-98187-614-1
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Неандертальцы: история несостоявшегося человечества: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Неандертальцы: история несостоявшегося человечества»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Автор рассматривает эти и многие другие вопросы, попутно суммируя и в доступной для неспециалистов форме излагая то, что известно сейчас о происхождении и эволюционной истории неандертальцев, их умственных и языковых способностях, материальной и зарождавшейся духовной культуре, о динамике их расселения и причинах вымирания. По каждой из перечисленных тем учтены наиболее интересные и важные сведения, имевшиеся в распоряжении палеоантропологии, археологии и смежных с ними наук на середину 2010 г.
Книга адресована всем, кого занимает древнейшее прошлое человечества — от академиков до студентов и школьников старших классов.
Неандертальцы: история несостоявшегося человечества — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Неандертальцы: история несостоявшегося человечества», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Всё перечисленное проделали и с ДНК неандертальцев. Результаты получились очень интересные. Однако прежде чем познакомиться с ними, нам следует, наверно, вспомнить некоторые азы биологии. Ну, хотя бы для того, чтобы у читателей, успевших несколько подзабыть школьный курс этой науки, не появилось подозрение, что их просто пытаются водить за нос с помощью непонятных слов.
Итак, ДНК. Эту аббревиатуру генетики придумали, чтобы не мучиться по сто раз на дню, выговаривая или набирая на клавиатуре компьютера слова «дезоксирибонуклеиновая кислота». Молекулы, или, точнее, макромолекулы ДНК — место хранения генетической информации, определяющей индивидуальные особенности и характер развития каждого организма и передаваемой от поколения к поколению. Каждая такая макромолекула — это цепочка, образованная двумя тяжами (нитями), спирально закрученными один относительно другого, а каждый тяж представляет собой последовательность тысяч и миллионов нуклеотидов. Эти нуклеотиды были бы похожи между собой, как близнецы, если бы не так называемые азотистые основания, входящие в их состав наряду с молекулой сахара (дезоксирибоза) и фосфата (фосфорная кислота) и представленные четырьмя разными типами. Два типа — гуанин и аденин — называются пуриновыми основаниями, а ещё два — тимин и цитозин — пиримидиновыми.
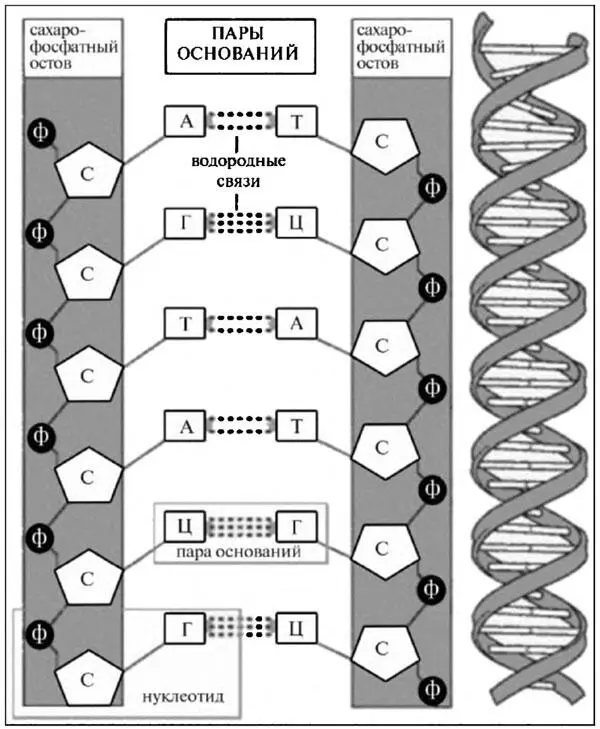
Рис. 3.14. Структура двойной спирали ДНК: Ф — фосфат, С — сахар, А — аденин, Т — тимин, Г — гуанин, Ц — цитозин
Молекулы сахара и фосфата образуют остов тяжей (нитей) ДНК, а основания находятся между тяжами и посредством слабых водородных связей соединяют между собой противолежащие нуклеотиды (рис. 3.14). При этом аденин может соединяться только с тимином, а гуанин только с цитозином. Последовательность этих парных оснований — шифр, в котором закодированы свойства белковых молекул и, в конечном счёте, свойства всего живого. Гены, т. е. сегменты ДНК, отвечающие за синтез разных белков, могут иметь длину от нескольких десятков до нескольких миллионов парных оснований.
Иногда вследствие слабости водородных связей при репликации ДНК происходят мутации, т. е. «перестановки» оснований, или, иными словами, изменения в порядке последовательности нуклеотидов. В большинстве своём такие мутации нейтральны по отношению к естественному отбору. Они не выбраковываются и не подхватываются им, поскольку не отражаются на приспособленности организмов, и благодаря этому обстоятельству скорость их накопления на молекулярном уровне в целом постоянна. Следовательно, если для истории изучаемой группы организмов имеются более или менее чёткие и надёжно датированные палеонтологические реперы (точки отсчёта), то эту скорость можно рассчитать. Например, для гоминид, а иногда и для всех обезьян вообще в качестве основы расчётов используется генетическое расстояние (т. е. количество различий в последовательности нуклеотидов в ДНК) между современными людьми и шимпанзе, эволюционные пути которых, судя по ископаемым находкам, разошлись около 6 млн. лет назад.
Зная скорость накопления мутаций и генетическое расстояние между разными особями или таксонами (т. е. видами, родами и т. д.), можно не только судить о близости их родства, но и рассчитывать время дивергенции, расхождения от общего предка, применяя для этого упоминавшийся уже выше метод молекулярных часов. Обычно для таких расчётов используют ДНК из митохондрий клеток (мтДНК), которая, в отличие от ДНК, содержащейся в ядрах, представлена в каждой клетке сотнями и тысячами идентичных копий, наследуется только по женской линии и к тому же характеризуется более высокими темпами накопления мутаций.
На основании изучения митохондриальной ДНК (мтДНК) были получены и первые сведения о генетике неандертальцев. Сначала, в 1997 г., удалось выделить пригодный для анализа фрагмент мтДНК из одной из костей, найденных в 1856 г. в гроте Фельдгофер, и сравнить его с аналогичным участком ДНК современного человека. Следующими «поставщиками» ископаемого генетического материала стали кости из пещер Виндия (Хорватия) и Мезмайская (Северный Кавказ, Россия). К настоящему времени генетические данные имеются уже примерно по 15 неандертальцам, жившим или погребённым в разное время в пещерах Испании (Эль Сидрон), Франции (Ля Шапелль-о-Сен, Ле Роше де Вилленёв), Бельгии (Анжи, Складина), Италии (Монте Лессини), Узбекистана (Тешик-Таш) и Южной Сибири (пещера Окладникова).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Неандертальцы: история несостоявшегося человечества»
Представляем Вашему вниманию похожие книги на «Неандертальцы: история несостоявшегося человечества» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.








Обсуждение, отзывы о книге «Неандертальцы: история несостоявшегося человечества» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.