Леонид Вишняцкий - Неандертальцы - история несостоявшегося человечества
Здесь есть возможность читать онлайн «Леонид Вишняцкий - Неандертальцы - история несостоявшегося человечества» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Санкт Петербург, Год выпуска: 2010, ISBN: 2010, Издательство: Нестор-История, Жанр: История, Биология, Прочая научная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Неандертальцы: история несостоявшегося человечества
- Автор:
- Издательство:Нестор-История
- Жанр:
- Год:2010
- Город:Санкт Петербург
- ISBN:978-5-98187-614-1
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Неандертальцы: история несостоявшегося человечества: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Неандертальцы: история несостоявшегося человечества»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Автор рассматривает эти и многие другие вопросы, попутно суммируя и в доступной для неспециалистов форме излагая то, что известно сейчас о происхождении и эволюционной истории неандертальцев, их умственных и языковых способностях, материальной и зарождавшейся духовной культуре, о динамике их расселения и причинах вымирания. По каждой из перечисленных тем учтены наиболее интересные и важные сведения, имевшиеся в распоряжении палеоантропологии, археологии и смежных с ними наук на середину 2010 г.
Книга адресована всем, кого занимает древнейшее прошлое человечества — от академиков до студентов и школьников старших классов.
Неандертальцы: история несостоявшегося человечества — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Неандертальцы: история несостоявшегося человечества», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
В составе семейства гоминид различают сейчас до семи разных родов и более двадцати видов. В то же время, по мнению целого ряда исследователей, обе эти цифры сильно завышены, и многие из традиционно выделяемых видов должны быть объединены между собой, а некоторые из наиболее ранних родов вообще исключены из числа близких родственников человека и переведены в предки шимпанзе, горилл или каких-то вымерших человекообразных обезьян. Преобладающие в настоящее время представления о родовом и видовом составе семейства гоминид, а также его эволюционной истории суммированы на рисунках 3.1 и 3.2 и в таблице 3.1.
За последние пятнадцать лет удалось существенно продвинуться в решении вопроса о времени рождения семейства гоминид. Произошло это не только благодаря новым палеонтологическим находкам, но и вследствие развития биомолекулярных методов датирования филогенетических событий. Принцип «молекулярных часов», лежащий в основе этих методов, отчасти сродни тому, на котором базируются радиоизотопные способы датирования. Если в последних в качестве основы расчётов используется примерно одинаковая для больших промежутков времени скорость распада радиоактивных элементов (например, C 14— радиоактивного изотопа углерода), то в первых аналогичную роль играют мутации, ведущие к изменению нуклеотидных последовательностей в ДНК или аминокислотных последовательностей в белках. Предполагается, что эти мутации распределяются во времени (конечно, речь о достаточно длительных его отрезках) более или менее равномерно. Если это так, то, сравнивая строение гомологичных белков или участков ДНК у разных групп организмов, можно судить о степени их родства (чем оно ближе, тем меньше должно быть различий), а при известной скорости накопления мутаций (её можно рассчитать, сравнивая ДНК тех видов, время расхождения которых установлено по надёжно датированным ископаемым останкам) даже и о древности дивергенции (расхождения) от общего предка.
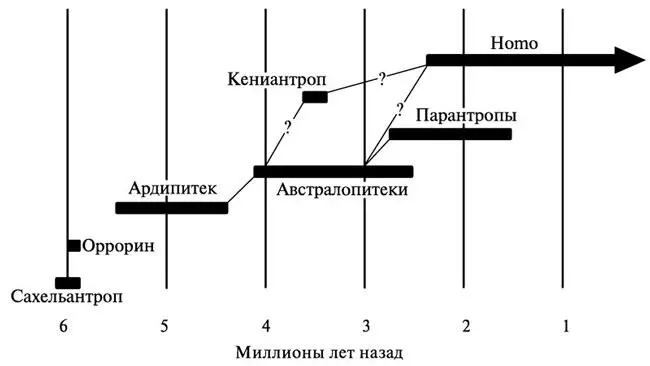
Рис. 3.1. Роды семейства гоминид на хронологической шкале. Показаны также их возможные генеалогические связи
Гипотеза «молекулярных часов» была сформулирована американскими биохимиками Э. Цукеркандлем и Л. Полингом в 1962 г. и почти сразу же стала широко использоваться для определения времени дивергенции разных групп животных, включая и приматов. Первая попытка применить её к изучению филогенетической истории человека и человекообразных обезьян была предпринята ещё в 1967 г., и с тех пор исследования такого рода продолжались, приобретая всё больший размах. Их результаты вкупе с датировками, полученными для ряда ключевых палеонтологических находок, говорят о том, что эволюционные пути наших предков и предков шимпанзе, ближайших родственников человека в современном животном мире, окончательно разошлись где-то в интервале от 8 до 5 млн. лет назад. Ископаемые материалы, имеющиеся для этого периода, заставляют думать, что прародиной гоминид, скорее всего, были северные районы Восточной и, возможно, Центральной Африки (хотя последнее менее вероятно). Именно оттуда происходят костные останки сахельантропа, оррорина и ардипитека — существ, рассматриваемых сейчас большинством исследователей в качестве древнейших представителей нашей эволюционной линии после её отделения от всех остальных гоминоидов. На смену им около 4 млн. лет назад пришли австралопитеки, которые, кроме двух названных выше регионов, освоили также и Южную Африку [57]. В настоящее время выделяют до девяти видов австралопитеков, и среди этих видов есть несколько кандидатов в наши родоначальники. Вполне возможно, что в будущем появятся ещё более достойные претенденты на эту роль — либо в лице какого-то неописанного пока вида австралопитеков [58], либо в лице представителей другого рода гоминид.
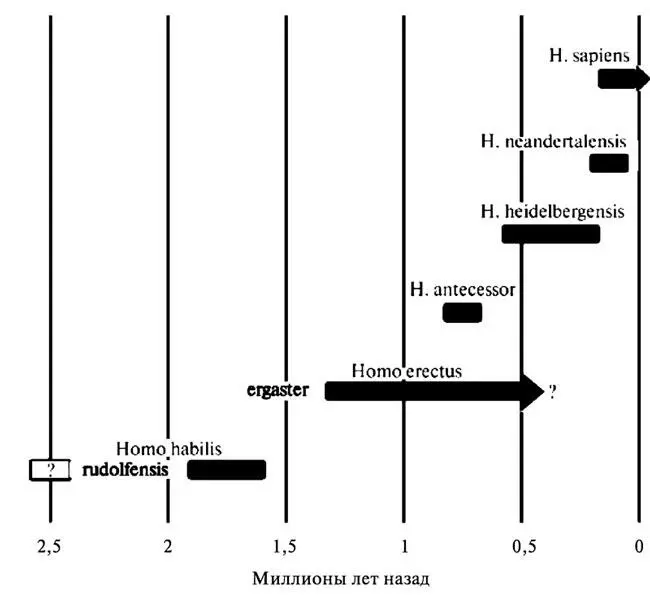
Рис. 3.2. Основные виды рода Homo на хронологической шкале
Таблица 3.1: Родовой и видовой состав семейства гоминид| Таксон | Когда впервые выделен | Область распространения находок | Датировки (млн. лет назад) |
|---|---|---|---|
| Sahelanthropus tchadensis | 2002 | Центральная Африка | 7,0 |
| Orrorin tugenensis | 2001 | Восточная Африка | 6,0 |
| Ardipithecus ramidus * | 1994 | Восточная Африка | 4,4 |
| Ardipithecus kadabba ** | 2004 | Восточная Африка | 5,8/5,2 |
| Australopithecus anamensis | 1995 | Восточная Африка | 4,2–3,9 |
| Australopithecus afarensis | 1978 | Восточная Африка | 3,9–3,0 |
| Australopithecus bahrelghazali | 1996 | Центральная Африка | 3,5/3,0 |
| Kenyanthropus platyops | 2001 | Восточная Африка | 3,5 |
| Australopithecus africanus | 1925 | Южная Африка | 3,0–2,4 |
| Australopithecus garhi | 1999 | Восточная Африка | 2,5 |
| Australopithecus sediba | 2010 | Южная Африка | 2,0–1,8 |
| Paranthropus aethiopicus *** | 1968 | Восточная Африка | 2,7–2,2 |
| Paranthropus robustus | 1938 | Южная Африка | 1,9–1,4 |
| Paranthropus boisei **** | 1959 | Восточная Африка | 2,3–1,2 |
| Homo habilis | 1964 | Восточная и Южная (?) Африка | 2,4–1,4 |
| Homo rudolfensis ***** | 1986 | Восточная Африка | 2,4 |
| Homo ergaster | 1975 | Восточная и Южная (?) Африка, Закавказье | 1,9–1,6 |
| Homo erectus ****** | 1894 | Африка, Восточная и Юго-Восточная Азия, юг Европы | 1,6–0,6 |
| Homo antecessor | 1997 | Западная Европа | 0,8 |
| Homo heidelbergensis | 1908 | Африка, Азия, Европа | 0,6–0,2 |
| Homo neanderthalensis | 1864 | Европа, Западная и Центральная Азия | 0,2–0,03 |
| Homo floresiensis | 2004 | Юго-Восточная Азия | 0,03–0,018 |
| Homo sapiens | 1758 | Повсеместно | 0,2–0 |
* Первоначально был описан под названием Australopithecus ramidus.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Неандертальцы: история несостоявшегося человечества»
Представляем Вашему вниманию похожие книги на «Неандертальцы: история несостоявшегося человечества» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.








Обсуждение, отзывы о книге «Неандертальцы: история несостоявшегося человечества» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.