Гдаль Оксенгендлер - Яды и противоядия
Здесь есть возможность читать онлайн «Гдаль Оксенгендлер - Яды и противоядия» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Ленинград, Год выпуска: 1982, Издательство: Наука, Жанр: Химия, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Яды и противоядия
- Автор:
- Издательство:Наука
- Жанр:
- Год:1982
- Город:Ленинград
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Яды и противоядия: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Яды и противоядия»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Яды и противоядия — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Яды и противоядия», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Холиеэстераза и ацетилхолин
Согласно существующим представлениям, в основе механизма действия ФОС лежит избирательное торможение ими фермента ацетилхолинэстеразы, или просто холинэстеразы, которая катализирует гидролиз ацетилхолина-химического передатчика (медиатора) нервного возбуждения. Различают 2 типа холинэстеразы: истинную, содержащуюся преимущественно в тканях нервной системы, в скелетной мускулатуре, а также в эритроцитах, и ложную, содержащуюся главным образом в плазме крови, печени и некоторых других органах. Собственно ацетилхолинэстеразой является истинная, или специфическая, холинэстераза, так как только она гидролизует названный медиатор. И именно ее в дальнейшем мы будем обозначать термином «холинэстераза». Поскольку фермент и медиатор являются необходимыми химическими компонентами передачи нервных импульсов в синапсах — контактах между двумя нейронами или окончаниями нейрона и рецепторной клеткой, следует более подробно остановиться на их биохимической роли.
Ацетилхолин синтезируется из спирта холина и ацетилкоэнзима А [70] Ацетилкоэнзим А — соединенно уксусной кислоты с нуклеотпи дом, включающим несколько аминокислот и активную SН-группу. Отщепляя ацетат, идущий на построение молекулы ацотилхолина, он превращается в коэнзим А.
под влиянием фермента холинацетилазы в митохондриях нервных клеток и накапливается в окончаниях их отростков в виде пузырьков диаметром около 50 нм. Предполагается, что каждый такой пузырек содержит несколько тысяч молекул ацетилхолина. При этом в настоящее время принято различать ацетилхолин, готовый к секреции и расположенный в непосредственной близости от активной зоны, и ацетилхолин вне активной зоны, находящийся в состоянии равновесия с первым и не готовый к выделению в сипаптическую щель. Кроме того, имеется еще так называемый стабильный фонд ацетилхолина (до 15%), не освобождающегося даже в условиях блокады его синтеза. [71] Глебов Р. Н., Примаковский Г. Н, Функциональная биохимия синапсов. М.: Медицина, 1978.
Под воздействием нервного возбуждения и ионов Са 2+молекулы ацетилхолина переходят в синаптическую щель — пространство шириною 20–50 нм, отделяющее окончание нервного волокна (пресинаптическую мембрану) от иннервируемой клетки. На поверхности последней расположена постсинаптическая мембрана с холинорецепторами — специфическими белковыми структурами, способными взаимодействовать с ацетилхолином. Воздействие медиатора на холинорецептор приводит к деполяризации (снижению заряда), временному изменению проницаемости постсинаптической мембраны для положительно заряженных ионов Na +и проникновению их внутрь клетки, что в свою очередь выравнивает потенциал напряжения на ее поверхности (оболочке). [72] Согласно установившейся точке зрения, возникновение разности потенциалов между наружной и внутренней сторонами поверхностного сдоя клетки обусловлено неравномерным распределением ионов Na + и К + по обе стороны клеточной мембраны. При этом компенсирующий поток ионов К + , направленный в обратную сторону при воздействии медиатора на постсинантическую мембрану, несколько запаздывает, что приводит на короткое время к обеднению внешней поверхности клетки положительными ионами.
Это дает начало новому импульсу в нейроне следующей ступени или вызывает деятельность клеток того или иного органа: мышцы, железы и др. (рис 5). Фармакологические исследования выявили существенную разницу в свойствах холинорецепторов различных синапсов. Рецепторы одной группы, проявляющие избирательную чувствительность к мускарину (яду гриба мухомора), названы мускариночувствительными, или М-холинорецепторами; они представлены главным образом в гладких мышцах глаз, бронхов, желудочно-кишечного тракта, в клетках потовых и пищеварительных желез, в сердечной мышце. Холинорецепторы второй группы возбуждаются малыми дозами никотина и поэтому названы никотино-чувствительными, или Н-холинорецепторами. К ним относятся рецепторы вегетативных ганглиев, скелетных мышц, мозгового слоя надпочечниковых желез, центральной нервной системы.
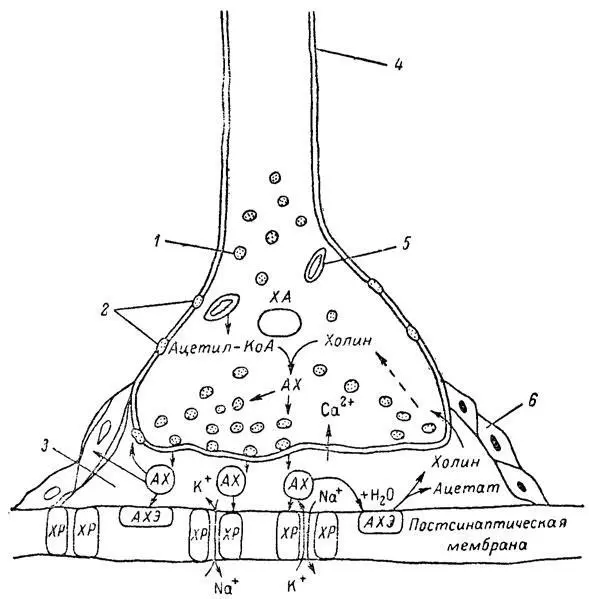
Молекулы ацетилхолина, выполнившие свою медиаторную функцию, должны быть немедленно инактивированы, в противном случае будет нарушена дискретность в проведении нервного импульса и проявится избыточная функция холинорецептора. Именно это осуществляет холинэстераза, мгновенно гидролизующая ацетилхолин. Каталитическая активность холинэстеразы превышает почти все известные ферменты: по разным данным, время расщепления одной молекулы ацетилхолина составляет около одной миллисекунды, что соизмеримо со скоростью передачи нервного импульса. Осуществление столь мощного каталитического эффекта обеспечивается наличием в молекуле холинэстеразы определенных участков (активных центров), обладающих исключительно хорошо выраженной реакционной способностью по отношению к ацетилхолину. [73] Розенгарт В. И. Холинэстеразы. Функциональная роль и клиническое значение. — В кн.: Проблемы медицинской химии. М.: Медицина, 1973, с. 66–104.
Будучи простым белком (протеином), состоящим только из одних аминокислот, молекула холинэстеразы, как теперь выяснено, исходя из ее молекулярной массы, содержит от 30 до 50 таких активных центров.
Интервал:
Закладка:
Похожие книги на «Яды и противоядия»
Представляем Вашему вниманию похожие книги на «Яды и противоядия» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «Яды и противоядия» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.