Александр Солодков - Физиология человека. Общая. Спортивная. Возрастная
Здесь есть возможность читать онлайн «Александр Солодков - Физиология человека. Общая. Спортивная. Возрастная» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2016, ISBN: 2016, Издательство: Литагент Спорт, Жанр: Биология, Детская образовательная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Физиология человека. Общая. Спортивная. Возрастная
- Автор:
- Издательство:Литагент Спорт
- Жанр:
- Год:2016
- Город:Москва
- ISBN:978-5-906839-67-1
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Физиология человека. Общая. Спортивная. Возрастная: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Физиология человека. Общая. Спортивная. Возрастная»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Для студентов, аспирантов, научных сотрудников, преподавателей, тренеров и врачей, работающих в области физической культуры.
Физиология человека. Общая. Спортивная. Возрастная — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Физиология человека. Общая. Спортивная. Возрастная», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Гомеостаз представляет собой не статическое явление, а динамическое равновесие. Способность сохранять гомеостаз в условиях постоянного обмена веществ и значительных колебаний факторов внешней среды обеспечивается комплексом регуляторных функций организма. Эти регуляторные процессы поддержания динамического равновесия получили название гомеокинеза.
Степень сдвига показателей гомеостаза при существенных колебаниях условий внешней среды или при тяжелой работе у большинства людей очень невелика. Например, длительное изменение pH крови всего на 0,1–0,2 может привести к смертельному исходу. Однако в общей популяции имеются отдельные индивиды, обладающие способностью переносить гораздо большие сдвиги показателей внутренней среды. У высококвалифицированных спортсменов-бегунов в результате большого поступления молочной кислоты из скелетных мышц в кровь во время бега на средние и длинные дистанции pH крови может снижаться до величин 7,0 и даже 6,9. Лишь несколько человек в мире оказались способными подняться на высоту порядка 8800 м над уровнем моря (на вершину Эвереста) без кислородного прибора, т. е. существовать и двигаться в условиях крайнего недостатка кислорода в воздухе и, соответственно, в тканях организма. Эта способность определяется врожденными особенностями человека – так называемой генетической нормой реакции, которая даже для достаточно постоянных функциональных показателей организма имеет широкие индивидуальные различия.
2.5. Возникновение возбуждения и его проведение
2.5.1. Мембранные потенциалы
Мембрана клетки состоит из двойного слоя молекул липидов, повернутых «головками» наружу, а «хвостами» друг к другу. Между ними свободно плавают глыбы белковых молекул. Некоторые из них пронизывают мембрану насквозь. В части таких белков имеются особые поры или ионные каналы, через которые могут проходить ионы, участвующие в образовании мембранных потенциалов (рис. 1, А).
В возникновении и поддержании мембранного потенциала покоя основную роль играют два специальных белка. Один из них выполняет роль особого натрий-калиевого насоса, который за счет энергии АТФ активно перекачивает натрий из клетки наружу, а калий внутрь клетки. В результате концентрация ионов калия становится внутри клетки выше, чем в омывающей клетку жидкости, а ионов натрия – выше снаружи.
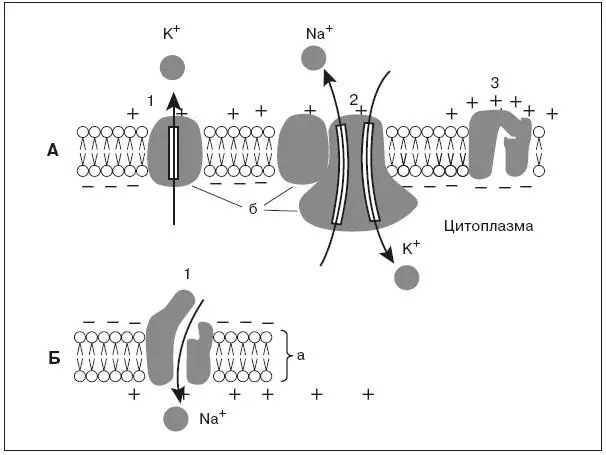
Рис. 1. Мембрана возбудимых клеток в покое (А) и при возбуждении (Б):
а – двойной слой липидов, б – белки мембраны;
А – каналы «утечки калия» (1), «натрий-калиевый насос» (2) и закрытый в покое натриевый канал (3); Б – открытый при возбуждении натриевый канал (1), вхождение ионов натрия в клетку и смена зарядов на наружной и внутренней стороне мембраны
Второй белок служит каналом утечки калия, через который ионы калия в силу диффузии стремятся выйти из клетки, где они содержатся в избытке. Ионы калия, выходя из клетки, создают положительный заряд на наружной поверхности мембраны. В результате внутренняя поверхность мембраны оказывается заряженной отрицательно по отношению к наружной. Таким образом, мембрана в состоянии покоя поляризована, т. е. имеется определенная разность потенциалов по обе стороны мембраны, называемая потенциалом покоя. Она равна для нейрона примерно минус 70 мВ, для мышечного волокна – минус 90 мВ. Измеряют мембранный потенциал покоя, вводя тонкий кончик микроэлектрода внутрь клетки, а второй электрод помещая в окружающую жидкость. В момент прокола мембраны и вхождения микроэлектрода внутрь клетки на экране осциллографа наблюдают смещение луча, пропорциональное величине потенциала покоя.
В основе возбуждения нервных и мышечных клеток лежит повышение проницаемости мембраны для ионов натрия – открывание натриевых каналов. Внешнее раздражение вызывает перемещение заряженных частиц внутри мембраны и уменьшение исходной разности потенциалов по обе стороны, или деполяризацию мембраны. Небольшие величины деполяризации приводят к открыванию части натриевых каналов и незначительному проникновению натрия внутрь клетки. Эти реакции являются подпороговыми и вызывают лишь местные (локальные) изменения.
При увеличении раздражения изменения мембранного потенциала достигают порога возбудимости, или критического уровня деполяризации, – около 20 мВ, при этом величина потенциала покоя снижается примерно до минус 50 мВ. В результате открывается значительная часть натриевых каналов. Происходит лавинообразное вхождение ионов натрия внутрь клетки, вызывающее резкое изменение мембранного потенциала, которое регистрируется в виде потенциала действия. Внутренняя сторона мембраны в месте возбуждения оказывается заряженной положительно, а внешняя – отрицательно (рис. 1,Б).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Физиология человека. Общая. Спортивная. Возрастная»
Представляем Вашему вниманию похожие книги на «Физиология человека. Общая. Спортивная. Возрастная» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «Физиология человека. Общая. Спортивная. Возрастная» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.