Array Сборник статей - Мозговой трест. 40 ведущих нейробиологов – о том, что мы знаем и чего не знаем о мозге
Здесь есть возможность читать онлайн «Array Сборник статей - Мозговой трест. 40 ведущих нейробиологов – о том, что мы знаем и чего не знаем о мозге» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2021, ISBN: 2021, Жанр: Биология, Прочая научная литература, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Мозговой трест. 40 ведущих нейробиологов – о том, что мы знаем и чего не знаем о мозге
- Автор:
- Жанр:
- Год:2021
- Город:Москва
- ISBN:978-5-00131-420-2
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Мозговой трест. 40 ведущих нейробиологов – о том, что мы знаем и чего не знаем о мозге: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Мозговой трест. 40 ведущих нейробиологов – о том, что мы знаем и чего не знаем о мозге»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Авторы описывают самые удивительные особенности мозга, честно объясняя, что известно, а что пока неизвестно ученым о работе нервной системы. Книга увлечет всех, кто интересуется наукой о мозге.
Мозговой трест. 40 ведущих нейробиологов – о том, что мы знаем и чего не знаем о мозге — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Мозговой трест. 40 ведущих нейробиологов – о том, что мы знаем и чего не знаем о мозге», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
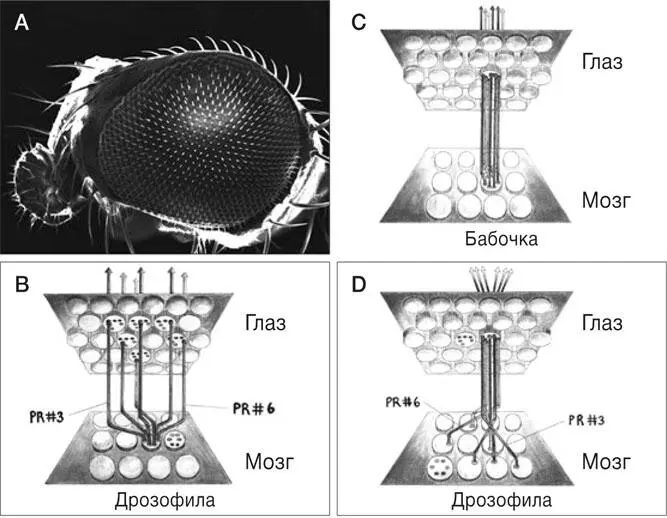
РИС. 2.Нейронные связи в глазу насекомого. (А) Поверхность глаза дрозофилы состоит из приблизительно 800 омматидиев. Обратите внимание на выпуклую поверхность каждого омматидия: это линза, которая фокусирует свет на фоторецепторах. (В) Схема связей между глазом и мозгом дрозофилы. Связи шести разных фоторецепторов (PR) глаза, воспринимающих свет одного направления (стрелки вверху) и расположенных в соседних омматидиях; аксоны сложными путями подходят к одной и той же мишени (патрону оптического ганглия) в мозге. Аксоны двух из шести таких фоторецепторов обозначены как PR#3 и PR#6. (C) Схема нейронных связей глаза и мозга у бабочки. В отличие от дрозофилы у бабочки фоторецепторы, воспринимающие свет одного направления (стрелки вверху), расположены в одном омматидии. Их аксоны тянутся напрямую к одному патрону – эта схема гораздо проще, чем у дрозофилы. (D) Схема связей между глазом и мозгом у дрозофилы. Все фоторецепторы одного омматидия воспринимают свет разных направлений (стрелки вверху), а их аксоны тянутся к шести разным патронам ламины
Адаптация заключается в том, что шесть фоторецепторов, расположенных в разных, но соседних омматидиях, воспринимают свет одного направления (рис. 2B, параллельные стрелки), а шесть фоторецепторов одного омматидия воспринимают свет разных направлений (рис. 2D, расходящиеся стрелки). Каким же образом аксоны фоторецепторов, воспринимающих свет одного направления и расположенных в соседних омматидиях, тянутся к одному, тому самому патрону ламины (рис. 2B)? [24]В отличие от глаза бабочки в глазу дрозофилы это не может осуществляться за счет того, что все фоторецепторы омматидия просто тянут свои аксоны к соответствующему оптическому патрону (сравните рис. 2В и 2С), и это усложняет задачу организации такого рода связей. На рис. 2В показана схема лишь одного набора из шести фоторецепторов в соседних омматидиях, соединяющихся с одним патроном ламины в мозге дрозофилы, однако нужно понимать, что таким сложным образом одновременно распределяются все 5000 аксонов фоторецепторов всех 800 омматидиев глаза дрозофилы – такой хореографии могли бы позавидовать даже постановщики танцевального шоу в перерыве Суперкубка. «Индивидуальные метки» для каждого соединения фоторецептора с ламиной едва ли помогли бы аксонам не сплестись в запутанный клубок, протягиваясь к своим патронам.
Петер Робин Хизингер и его коллеги исследовали механизмы формирования нейронных связей в глазу дрозофилы тем способом, который одобрил бы Рамон-и-Кахаль: они просто наблюдали [25]. Воспользовавшись возможностями современных методов микроскопии, которые позволяют получать изображения отдельных аксонов фоторецепторов и их конусов роста, тянущихся к мишеням, ученые вывели простые правила, по которым образуются связи в глазу дрозофилы. Микроскоп – это прибор, который позволяет наблюдателю заглянуть внутрь живой ткани и во всех подробностях разглядеть строение отдельных нейронов, аксонов и их конусов роста [26]. Эксперимент заключался в наблюдении за аксонами фоторецепторов и их конусами роста на протяжении около 30 часов: столько времени требуется на рост аксона от глаза к головному мозгу у дрозофилы. Тщательный анализ интервальной съемки позволил однозначно идентифицировать каждый из шести фоторецепторов в каждом из 800 омматидиев глаза дрозофилы.
Главный результат наблюдений: в каждом из шести фоторецепторов в любом отдельном омматидии действуют разные модели роста аксона после его контакта с ламиной. Например, аксон от фоторецептора #3 всегда сначала достигает ламины «по прямой», а затем тянется «на юго-восток» с постоянной скоростью (рис. 2B и 2D). И эта модель роста одинакова для всех фоторецепторов #3, расположенных в разных омматидиях глаза. Аксон фоторецептора #6 достигает ламины «по прямой», а затем меняет направление и растет «на запад»; скорость его роста тоже постоянна, но отличается от скорости роста аксона фоторецептора #3. Аналогичным образом ведут себя остальные четыре фоторецептора. Если сравнить направления роста аксонов фоторецепторов #1–6 из разных омматидиев, то выяснится, что они идентичны. Получается, что в каждый из шести фоторецепторов встроена уникальная программа роста, которая выполняется одинаково в каждом омматидии и определяет правило, по которому собирается сложная схема соединений нейронов в глазу дрозофилы. Если правило выполняется и все шесть аксонов фоторецепторов растут в заданном направлении с заданной скоростью, это дает удивительный результат: шесть конусов роста аксонов шести фоторецепторов, расположенных в соседних омматидиях и воспринимающих свет одного направления, в одно и то же время сходятся в одном патроне оптического ганглия, и рост останавливается (рис. 2В). Из этого следует второе правило: рост аксонов прекращается только после того, как конусы роста всех шести аксонов фоторецепторов достигают ламины и соприкасаются там друг с другом, – и никак не раньше. Таким образом, аксоны фоторецепторов, воспринимающих свет одного направления, находят свой путь через сеть тесно переплетенных аксонов других нейронов и их конусов роста, и процесс не прерывается, если они соприкасаются с конусами роста аксонов фоторецепторов, воспринимающих свет иного направления. Механизм формирования связей между фоторецепторами и мозгом крайне точен, а ошибки редки. Каждый патрон оптического ганглия получает сигналы только от фоторецепторов, воспринимающих свет одного направления. Это значительно упрощает обработку визуальных сигналов в отделах мозга более высокого уровня – ведь информация о направлении анализируется еще на уровне патрона ламины, первой «ретрансляторной станции» в зрительной системе, сразу после восприятия света фоторецептором. Компьютерное моделирование, выполненное Хизингером и его коллегами, показало, что для формирования нужной связи достаточно одновременной встречи шести конусов роста аксонов фоторецепторов; никаких меток в патроне ламины не требуется. Таким образом, трудноразрешимая, на первый взгляд, задача формирования сложной схемы соединения 5000 аксонов фоторецепторов, необходимой для суперпозиции нейронов, решается при помощи всего шести разных программ роста аксонов фоторецепторов в процессе развития глаза. Очевидно, чтобы направить каждый из приблизительно 5000 аксонов фоторецепторов к своей мишени, не требуется никаких сложных направляющих меток.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Мозговой трест. 40 ведущих нейробиологов – о том, что мы знаем и чего не знаем о мозге»
Представляем Вашему вниманию похожие книги на «Мозговой трест. 40 ведущих нейробиологов – о том, что мы знаем и чего не знаем о мозге» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Мозговой трест. 40 ведущих нейробиологов – о том, что мы знаем и чего не знаем о мозге» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.