Сергей Ястребов - От атомов к древу. Введение в современную науку о жизни
Здесь есть возможность читать онлайн «Сергей Ястребов - От атомов к древу. Введение в современную науку о жизни» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: Москва, Год выпуска: 2018, ISBN: 2018, Издательство: Альпина нон-фикшн, Жанр: Биология, sci_biochem, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:От атомов к древу. Введение в современную науку о жизни
- Автор:
- Издательство:Альпина нон-фикшн
- Жанр:
- Год:2018
- Город:Москва
- ISBN:978-5-9614-5286-0
- Рейтинг книги:5 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
От атомов к древу. Введение в современную науку о жизни: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «От атомов к древу. Введение в современную науку о жизни»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Рекомендуется широкому кругу читателей, всерьез интересующихся современной биологией.
От атомов к древу. Введение в современную науку о жизни — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «От атомов к древу. Введение в современную науку о жизни», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
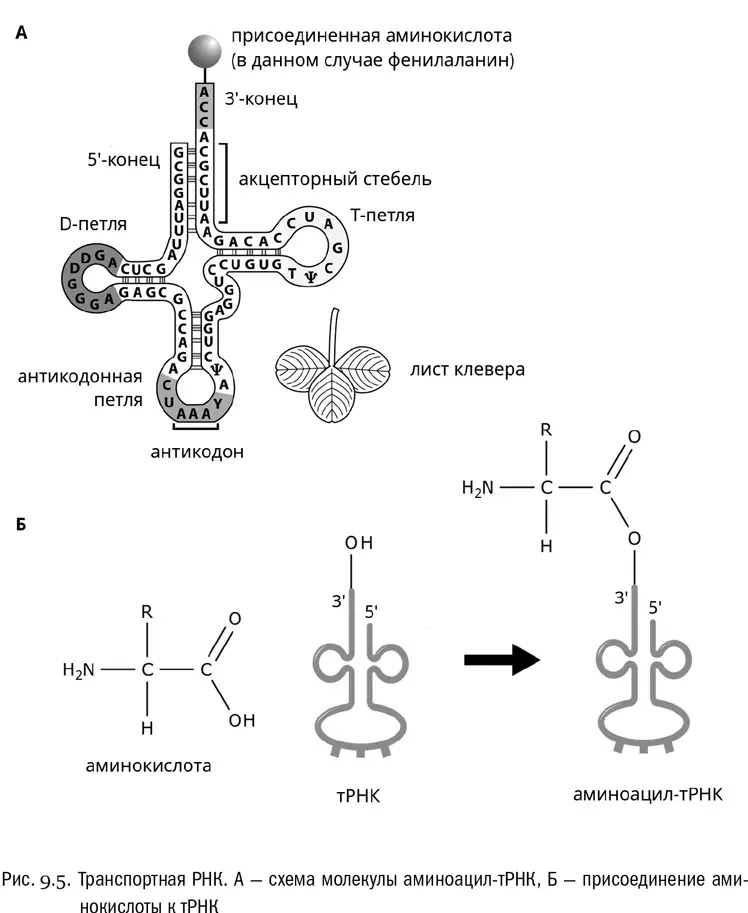
Само присоединение аминокислоты выглядит так (см. рис. 9.5Б). Фермент аминоацил-тРНК-синтетаза (он же просто кодаза) сшивает с выделением воды 3'-гидроксил концевого аденозина тРНК и карбоксильную группу аминокислоты. Последняя тем самым временно превращается в ковалентно связанный с рибозой остаток R−CH(NH 2)−CO−, который называется аминоацилом. Также в этой реакции участвует АТФ, который расщепляется в ходе нее до АМФ. Но главный продукт реакции — это аминоацил-тРНК, то есть молекула транспортной РНК с висящей на «черешке клеверного листа» ковалентно пришитой аминокислотой, временно превратившейся в аминоацил.
На вершине самой дальней от «черешка» петли тРНК всегда находится антикодон — нуклеотидный триплет, комплементарный кодону той самой аминокислоты, которую данная тРНК переносит. Например, если аминокислота фенилаланин кодируется кодоном УУУ, то соответствующая транспортная РНК несет антикодон ААА (поскольку, как мы знаем, урацил комплементарен аденину). Чтобы соединение произошло правильно, активный центр кодазы должен одновременно распознать и молекулу тРНК, и аминокислоту — не какую угодно, а ту, которая кодируется триплетом, комплементарным антикодону этой тРНК. Это сложная задача, но кодаза с ней обычно справляется.
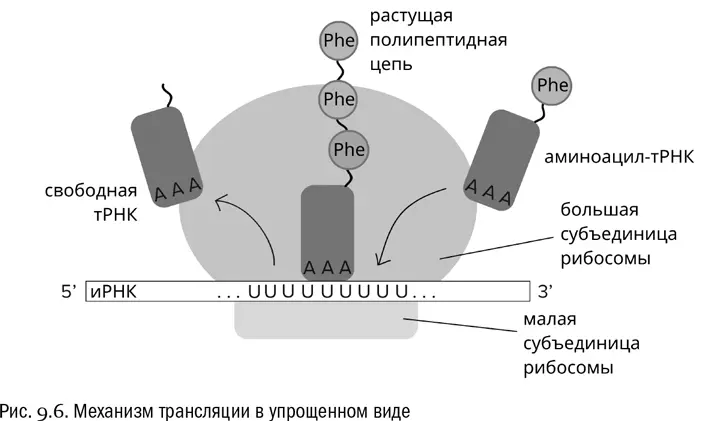
Дальше вступает в действие простой механизм (см. рис. 9.6). Во время трансляции любая проплывающая мимо аминоацил-тРНК может чисто случайно столкнуться с тем кодоном иРНК, который в данный момент находится в активном центре рибосомы. Но свяжется она с ним только в том случае, если ее антикодон будет этому кодону комплементарен. Тогда рибосома отрежет аминокислоту от тРНК, присоединит ее к растущей белковой цепочке, а сама продвинется по иРНК на один шаг вперед (в сторону 3'-конца). После чего цикл повторится.
Антикодон тРНК, связанной с рибомосой, всегда комплементарен кодону, находящемуся в данный момент в активном центре этой рибосомы. Иначе трансляция не пойдет. Добавим, что белок при трансляции всегда синтезируется от N-конца к C-концу, то есть от свободной аминогруппы к свободному карбоксилу, но не наоборот (см. главу 3). Именно поэтому аминокислотные последовательности белков в таком же порядке и записываются в базах данных.
Источниками энергии для трансляции (и для транскрипции тоже) служат нуклеозидтрифосфаты. Причем в данном случае это не столько уже знакомый нам АТФ, сколько гораздо менее распространенный ГТФ. Почему так? Ведь АТФ в клетке больше, а с точки зрения энергии выбор между АТФ и ГТФ не дает ни заметного проигрыша, ни заметного выигрыша, энергетическая «цена» реакций распада этих веществ примерно одна и та же.
Поставим вопрос иначе: почему вообще самой универсальной «энергетической валютой» стал АТФ, а не ГТФ, ЦТФ или УТФ? Простейшую подсказку можно получить, если взглянуть на формулы четырех азотистых оснований. Аденин — единственное из них, в котором нет ни одного атома кислорода. В молекулах гуанина, цитозина и урацила кислород есть. На древней Земле, где свободного кислорода в атмосфере было очень мало, аденин наверняка легче всего синтезировался, и, соответственно, адениновые нуклеотиды тоже. Клетки использовали тот химический субстрат, который был самым доступным. Возможно, как раз поэтому именно молекула АТФ стала универсальным переносчиком энергии [67] Здесь воспроизведена идея, которую высказал в сетевом обсуждении китайский биохимик Минь Чжоу: https://www.researchgate.net/post/Why_did_evolution_favor_ATP_and_not_GTP_TTP_or_CTP
.
Но универсальность АТФ имеет и свои минусы. Дело в том, что из-за особой важности этого вещества его концентрация (а точнее, соотношение концентраций [АТФ]/[АМФ]) очень жестко контролируется внутриклеточными регуляторными системами. В многоклеточном организме слишком резкое отклонение этого параметра от нормы может вызвать даже «самоубийство» целой клетки, так называемый апоптоз. На ГТФ этот контроль не распространяется, поэтому менять его концентрацию можно гораздо свободнее. Возможно, смысл «подключения» транскрипции и трансляции к ГТФ состоит как раз в том, чтобы сделать эти жизненно важные процессы автономными, снизив их зависимость от всего остального происходящего в клетке.
Открытие механизма трансляции тут же дало ученым превосходный ключ к расшифровке генетического кода. Например, что будет, если синтезировать искусственную иРНК, в которую из всех азотистых оснований входит только урацил, и поместить ее в обычный водный раствор, предварительно добавив туда рибосомы, полный набор аминоацил-тРНК и источники энергии? Оказалось, что в этом случае прямо в пробирке, без всякого участия живых клеток, может синтезироваться белок, состоящий из одной-единственной аминокислоты, а именно из фенилаланина. Этот эксперимент был реально поставлен в 1960 году, и в результате его был расшифрован первый кодон — УУУ. Это кодон фенилаланина [68] Вот описание этого опыта, которое в данном случае будет лучше любого пересказа своими словами: «В экспериментах с бесклеточной системой Маршалл Ниренберг и Генрих Маттэи, исследовавшие активность различных препаратов РНК в роли матриц для белкового синтеза, в качестве контроля использовали синтетическую полиуридиловую кислоту (poly U), рассчитывая, что она не будет проявлять существенной матричной активности. К своему большому удивлению, они обнаружили, что poly U достаточно эффективно направляет синтез полифенилаланина. Более того, полифенилаланин оказался единственным полипептидом, синтезируемым в присутствии poly U. Из этих наблюдений непосредственно вытекало, что триплет UUU служит кодоном для фенилаланина. Вскоре аналогичным образом было установлено, что poly C направляет синтез полипролина, а poly A — полилизина, то есть CCC является пролиновым кодоном, а AAA кодирует лизин. К счастью, использованная в этих экспериментах бесклеточная система содержала повышенную концентрацию ионов магния, при которой (как выяснилось в дальнейшем) инициация синтеза полипептидной цепи происходит и в отсутствие инициаторного кодона AUG. Только поэтому вышеупомянутые синтетические матрицы и удалось использовать для аномальной инициации трансляции. Так, отчасти благодаря счастливой случайности, удалось сделать первые шаги на пути к полной расшифровке генетического кода». ( Кайгер Д., Айала Ф. Современная генетика. — М.: Мир, 1987. Т. 2. С. 76.)
. Расшифровка всех остальных кодонов после этого была уже только делом биохимических опытов, пусть и непростых технически, но абсолютно прозрачных по смыслу. Завершить ее удалось всего за каких-то пять лет. К 1965 году генетический код был полностью взломан (cracked). Именно так это тогда называли в статьях, а еще чаще в разговорах, вполне в духе основоположника научного мировоззрения сэра Фрэнсиса Бэкона, некогда заявившего на весь мир, что знание — сила.
Интервал:
Закладка:
Похожие книги на «От атомов к древу. Введение в современную науку о жизни»
Представляем Вашему вниманию похожие книги на «От атомов к древу. Введение в современную науку о жизни» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «От атомов к древу. Введение в современную науку о жизни» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.