Дэвид Линден - Осязание. Чувство, которое делает нас людьми
Здесь есть возможность читать онлайн «Дэвид Линден - Осязание. Чувство, которое делает нас людьми» — ознакомительный отрывок электронной книги совершенно бесплатно, а после прочтения отрывка купить полную версию. В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Год выпуска: 2019, Жанр: Биология, sci_popular, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Осязание. Чувство, которое делает нас людьми
- Автор:
- Жанр:
- Год:2019
- ISBN:нет данных
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Осязание. Чувство, которое делает нас людьми: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Осязание. Чувство, которое делает нас людьми»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Профессор неврологии и известный популяризатор науки Дэвид Линден увлекательно и доступно – буквально «на пальцах»– объясняет, как работают сложные механизмы осязания, а заодно разбирает его многочисленные загадки. Почему перец кажется нам жгучим, а мята – холодной? Почему мы боимся щекотки, если нас щекочет кто-то другой, и не реагируем на нее, если пытаемся пощекотать себя сами? Что на самом деле происходит там, где чешется? Чем оргазм принципиально отличается от других мышечных спазмов, например чихания?
В книге Дэвида Линдена читатель найдет ответы на эти и многие другие вопросы.
Осязание. Чувство, которое делает нас людьми — читать онлайн ознакомительный отрывок
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Осязание. Чувство, которое делает нас людьми», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Зуд могут вызывать самые разные типы стимуляции кожи. Во многих случаях мы даже не обладаем пока пониманием молекулярных эффектов, вызывающих зуд. Для большинства стимулов зуда путь в мозг оказывается непрямым. Например, если кожа сильно натерта или проявляет местную реакцию на аллерген, включается воспалительный каскад (мы говорили о нем в главе 6 – см. рис.6.5). Молекулы, выделяемые иммунными клетками (например, гистамин из тучных клеток), могут поступать в гистаминовые рецепторы, расположенные на свободных окончаниях сенсорных нейронов в эпидермисе, и побуждать их испускать электрические импульсы (рис.7.2). В другом примере фрагмент естественного белка BAM8-22 поступает в другой рецептор на проводящих зуд нервных окончаниях кожи, который применительно к мышам называют MrgprC11, а к людям – hMrgprX1. Иногда происходит непосредственная активация рецептора зуда в окружающей среде. Например, хорошо известно, что противомалярийный препарат хлорохин вызывает зуд. Хлорохин непосредственно поступает в другой рецептор сенсорных нейронов, который называется MrgprA3. Отметим, что существует по меньшей мере три молекулярных сенсора, которые активируют нейроны, ответственные за распознавание зуда. И если некоторые активируются непосредственно сигналами окружающей среды, то большинство реагируют на химический сигнал-посредник в самом организме.
Если действительно существуют особые нейроны, отвечающие за зуд, то верны и следующие утверждения: 1) мы можем разрушать или подавлять эти нейроны и блокировать ощущение зуда, причем другие осязательные ощущения – боль и температура – останутся неизменными; 2) избирательная активация этих специализированных нейронов зуда должна вызывать ощущение зуда, но не боли и не других осязательных ощущений; 3) анатомическое распределение нервных окончаний отражает известное распределение ощущения зуда: они должны присутствовать в эпидермисе и во внешних слизистых мембранах, но отсутствовать в мышцах, связках, внутренних органах и т.д.
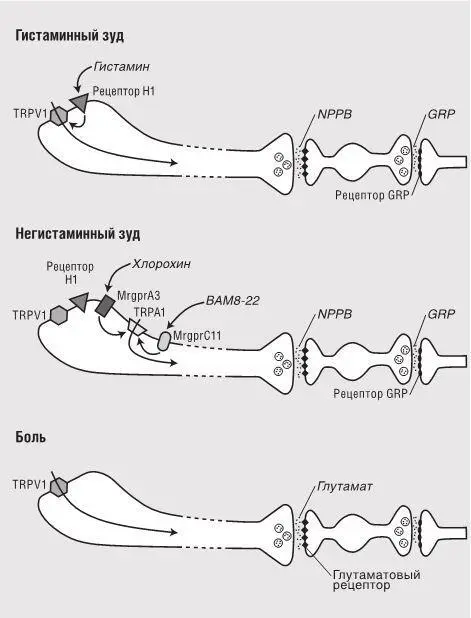
Рис.7.2. Два различных пути С-волокон при зуде в сопоставлении с болью. Нейромедиатор NPPB, по всей вероятности, специфичен для нейронов зуда. Напротив, нейроны боли выделяют глутамат, тем самым отправляя сигнал в нейроны заднего рога спинного мозга. Эти нейроны содержат рецепторы NPPB и, в свою очередь, выделяют редкий нейромедиатор GRP, сигнализируя им следующим нейронам в цепочке. Удаление нейронов с рецепторами GRP блокирует ощущение зуда, но не ощущение боли или легких прикосновений, что дает основание предположить, что эти два синаптических соединения специфичны именно для зуда [134] . Нейроны зуда можно разделить по меньшей мере на две категории: те, что содержат рецептор хлорохина MrgprA3 и в основном передают негистаминный зуд, и те, которые обладают только гистаминовым рецептором и отвечают за гистаминный зуд. Гистаминовые рецепторы возбуждают нервные окончания, открывая ионный канал TRPV1, а рецепторы хлорохина и BAM8-22 открывают ионный канал TRPA1. Эта диаграмма дает лишь общую схему. Скорее всего, существуют и другие популяции нейронов зуда, помимо показанных здесь. Кроме того, синаптические взаимодействия между потоками информации в спинном мозге на данный момент не вполне понятны
Один из подходов к определению потенциально специализированных нейронов – попытка выявить молекулу-нейромедиатор, используемую этими особыми нейронами для связи с соответствующими участками спинного мозга, и последующее удаление этой молекулы у мышей при помощи генетических манипуляций. Сантош Мишра и Марк Хун из Национального института здравоохранения так и поступили, сделав на основании опыта предположение о том, что нейротрансмиттер зуда – это молекула NPPB. [135]Выяснилось, что мышь-мутант без NPPB почти не испытывает зуда в ответ на множество стимулов, включая и гистамин, и хлорохин. А главное – мыши без NPPB нормально реагировали на боль, температуру и легкие прикосновения.
NPPB высвобождается аксонами сенсорных нейронов и передается таргетным нейронам заднего рога спинного мозга. У этих нейронов есть рецепторы, которые взаимодействуют с NPPB и распространяют электрические сигналы далее в головной мозг. После искусственного синтеза NPPB и впрыскивания этого вещества в спинной мозг мышей животные начали чесаться точно так же, как если бы испытывали зуд на коже в обычных условиях. После инъекции в спинной мозг особого токсина, который избирательно разрушает нейроны с рецепторами NPPB, мыши не отреагировали ни на применение зудящих стимулов, ни на впрыскивание NPPB в спинной мозг. Эти результаты позволяют предположить, что нейроны, использующие NPPB, специфичны именно для зуда. Если это верно, то избирательная активация этих нейронов должна вызывать зуд, но не боль и не ощущения легких прикосновений. На время создания этой книги отчетов о таких экспериментах еще не появилось, но, судя по всему, в некоторых лабораториях их пытаются провести.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Осязание. Чувство, которое делает нас людьми»
Представляем Вашему вниманию похожие книги на «Осязание. Чувство, которое делает нас людьми» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.












Обсуждение, отзывы о книге «Осязание. Чувство, которое делает нас людьми» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.