Александр Коган - Основы физиологии высшей нервной деятельности
Здесь есть возможность читать онлайн «Александр Коган - Основы физиологии высшей нервной деятельности» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Город: М., Год выпуска: 1988, ISBN: 1988, Издательство: Высшая школа, Жанр: Биология, Медицина, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Основы физиологии высшей нервной деятельности
- Автор:
- Издательство:Высшая школа
- Жанр:
- Год:1988
- Город:М.
- ISBN:5-06-001444-4
- Рейтинг книги:4 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Основы физиологии высшей нервной деятельности: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Основы физиологии высшей нервной деятельности»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Максимальная ширина картинки в этой fb2-книге — 600 пикселей, но большинство картинок имеют размер не более 460 пикселей.
Основы физиологии высшей нервной деятельности — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Основы физиологии высшей нервной деятельности», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:

Рис. 13. Синхронизация потенциалов моторной коры мозга кролика ритмическими вспышками света, совпадающими по частоте с электрокожным раздражением лапы при выработке двигательного оборонительного рефлекса. А — до выработки условного рефлекса; Б — после 150 сочетаний (по М.Н. Ливанову, К.Л. Полякову):
в каждой паре записей нижняя продолжает верхнюю, точками обозначены вспышки света, Ур — условно-рефлекторное движение лапы
Дальнейшие исследования показали, что реакция десинхронизации электрокортикограммы (ЭКоГ) человека и животных, так же как реакция усвоения ритма, может отражать многие свойства нервных механизмов условно-рефлекторной деятельности. Так, во время генерализации условных рефлексов эти реакции возникают на обширной территории коры, охватывая области проекций и условного и безусловного раздражителей, а по мере специализации условных рефлексов сосредоточиваются на более ограниченной зоне, большей частью в двигательной области коры.
В процессе образования временной связи реакции синхронизации и усвоения ритмов раздражителя вначале формируются в корковых проекциях сигнала, затем в зоне проекции подкрепления, при дальнейшем упрочении условного рефлекса идут на убыль в зоне проекции сигнала, но долго сохраняются в зоне проекции подкрепления. При этом усвоение ритма происходит сначала в верхних, а потом в нижних слоях коры. Парная работа полушарий в формировании временных связей проявляется в том, что по мере выработки условного рефлекса ритмы сигнального стимула усваиваются вначале структурами верхних слоев коры полушария, контралатерального стороне подкрепления, затем они появляются в его нижних слоях и во всех слоях ипсилатерального полушария, потом они уменьшаются и исчезают всюду, кроме нижних слоев полушария, контралатерального стороне подкрепления (рис. 14).
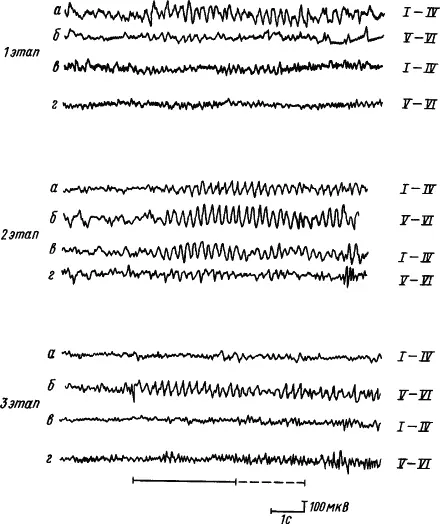
Рис. 14. Пространственная динамика усвоения ритма условного раздражителя в слоях симметричных пунктов коры больших полушарий головного мозга в процессе выработки условного рефлекса (по А.Б. Когану, Г.А. Кураеву):
I—VI — слои коры, а, б — запись ЭКоГ от полушария контралатерального относительно стороны подкрепления, в, г — то же, из ипсилатерального полушария; 1, 2, 3 — этапы выработки условного рефлекса ( 1 — первые сочетания, 2 — генерализованный условный рефлекс, 3 — упроченный специализированный), сплошной линией внизу обозначено действие условного стимула, пунктиром — безусловного
Разработка методов электроэнцефалографии с одновременной регистрацией ЭКоГ от десятков и сотен пунктов коры и использование электронно-вычислительных машин позволили выяснить роль синхронизации ритмов биоэлектрической активности в областях корковой проекции условного и безусловного раздражителей для образования временной связи между ними. Показано, например, что условные оборонительные движения лапы у кролика проявлялись тогда, когда коэффициент корреляции ритмов активности исследуемых корковых нейронов достигал достаточно высокого значения. Это подтверждено и для спонтанных движений. Функциональное значение пространственной синхронизации электрических потенциалов головного мозга было показано на многочисленных примерах. Показатели синхронизации также демонстрируют иррадиацию нервных процессов при начальной генерализации условного рефлекса, а по мере его закрепления и специализации нарастающую их концентрацию (рис. 15).
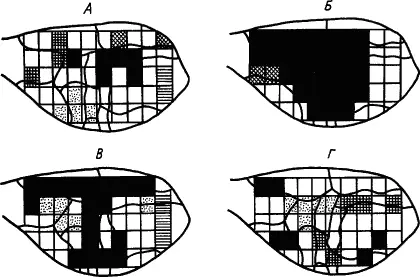
Рис. 15. Динамика синхронизации волн электрокортикограммы кролика в процессе выработки условного рефлекса. А — исходный фон; Б — после 12 сочетаний; В — после 60 сочетаний (появился условный рефлекс); Г — после 90 сочетаний (прочный условный рефлекс):
квадратами обозначены участки отведения; одинаково изображены участки, синхронность волн между которыми сохраняется не менее 70 % времени измерения
Многие сведения, существенные для понимания механизма образования временных связей, были получены при исследовании разных режимов условного раздражения и подкрепления, а также пространственных характеристик регистрируемой активности. Так, успех образования временной связи по показателям синхронизации усвоенных ритмов оказался зависимым главным образом от четырех факторов: 1) изоритмии сигнала и подкрепления; 2) оптимальности их частоты (для кролика 4–7 Гц); 3) отношения фаз (синфазность) и 4) достаточного объема нервного субстрата, вовлеченного в этот процесс. Отсюда сделано заключение, что наблюдаемая при выработке условных рефлексов синхронизация электрических потенциалов в разных структурах мозга отражает перестройку их лабильности, которая делает возможным движение нервных процессов между ними. Временная связь формируется на основе такого «проторения» пути синхронизированными ритмами биоэлектрической активности (М.Н. Ливанов, 1980).
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Основы физиологии высшей нервной деятельности»
Представляем Вашему вниманию похожие книги на «Основы физиологии высшей нервной деятельности» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.










Обсуждение, отзывы о книге «Основы физиологии высшей нервной деятельности» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.