Станислав Дробышевский - Шанс для приматов
Здесь есть возможность читать онлайн «Станислав Дробышевский - Шанс для приматов» весь текст электронной книги совершенно бесплатно (целиком полную версию без сокращений). В некоторых случаях можно слушать аудио, скачать через торрент в формате fb2 и присутствует краткое содержание. Издательство: сет. публикация, Жанр: Биология, на русском языке. Описание произведения, (предисловие) а так же отзывы посетителей доступны на портале библиотеки ЛибКат.

- Название:Шанс для приматов
- Автор:
- Издательство:сет. публикация
- Жанр:
- Год:неизвестен
- ISBN:нет данных
- Рейтинг книги:3 / 5. Голосов: 1
-
Избранное:Добавить в избранное
- Отзывы:
-
Ваша оценка:
Шанс для приматов: краткое содержание, описание и аннотация
Предлагаем к чтению аннотацию, описание, краткое содержание или предисловие (зависит от того, что написал сам автор книги «Шанс для приматов»). Если вы не нашли необходимую информацию о книге — напишите в комментариях, мы постараемся отыскать её.
Шанс для приматов — читать онлайн бесплатно полную книгу (весь текст) целиком
Ниже представлен текст книги, разбитый по страницам. Система сохранения места последней прочитанной страницы, позволяет с удобством читать онлайн бесплатно книгу «Шанс для приматов», без необходимости каждый раз заново искать на чём Вы остановились. Поставьте закладку, и сможете в любой момент перейти на страницу, на которой закончили чтение.
Интервал:
Закладка:
Другой пример быстрейшей специализации — летучие мыши Chiroptera. Рукокрылые, вероятно, имелись уже в верхнем мелу Южной Америки и верхнем палеоцене Франции и Германии (Gingerich, 1987; Hand et al. 1994; Hooker, 1996), а однозначные представители нижнего эоцена мало отличимы от современных, причём обнаружены сразу в десятках видов на всех континентах, включая Австралию.
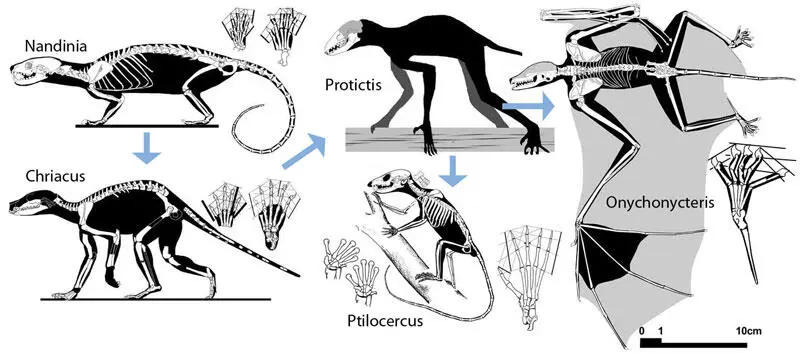
Происхождение рукокрылых
Замечательно, что зубы нижнеэоценовых летучих мышей почти идентичны зубам примитивных плацентарных типа Cimolestes и древнейших землеройковых, так что родство всех этих групп не представляет сомнения, что однозначно подтверждается данными генетики. Несмотря на то, что в генетико-кладистических схемах рукокрылые попадают в лавразиатериев Laurasiatheria, а приматы — в эуархантоглиресов Euarchontoglires, сходство двух этих групп всегда было очевидно всем систематикам, начиная с К. Линнея, и отразилось в создании группы "архонта" Archonta, объединяющей летучих мышей, приматов, тупайй и шерстокрылов. Сходство пращуров представителей "архонта" усиливалось древесным образом жизни предков летучих мышей и шерстокрылов и, как минимум, преадаптацией к нему у предков приматов и тупайй. Очевидно, потому и не удаётся выявить непосредственных нижнепалеоценовых или меловых предков летучих мышей, что их зубы неотличимы от зубов прочих примитивных зверей. Не исключено, что какие-то палеоценовые формы, известные лишь по зубам и считающиеся ныне приматами, плезиадаписовыми или какими-либо насекомоядными в широком смысле, при лучшей изученности окажутся примитивными летучими мышами. Пока у летучих мышей не было крыльев и эхолокации, мы считаем их "насекомоядными", когда же эти специализации появляются (судя по Onychonycteris finneyi, полёт возник раньше эхолокации (Simmons et al., 2008)), мы уже видим готовых рукокрылых.
Как и в случае с птицами и птерозаврами, машущий полёт летучих мышей возник очень быстро, и уловить момент его становления крайне трудно, для этого надо обладать невероятным везением.
Рукокрылые уникальны в том отношении, что первые этапы их эволюции отличались максимальными темпами, а последующие — крайне низкими (вернее, на уровне видо- и родообразования темпы были велики, но план строения уже на уровне семейств практически не изменился с нижнего эоцена); можно даже утверждать, что макроэволюция летучих мышей закончилась в то время, когда у приматов она только начиналась. Очевидно, что причиной этого стала адаптация к полёту. И без того небогатые заделы строения мозга первопредков были безнадёжно задавлены необходимостью облегчения веса; наглядно это выражается в быстром зарастании швов черепа, что было характерно уже для раннеэоценового Icaronycterys. О хватательной способности конечностей речь тоже не идёт, скорее уж о цеплятельной; нижнеэоценовый Onychonycteris имел когти на всех пальцах крыла, а остальные синхронные родственники — уже потеряли на двух или трёх.

Ископаемая летучая мышь Icaronycterys (ранний эоцен)
Правда, у рукокылых есть два существенных преимущества перед насекомоядными: они долго живут, а потому могут накопить богатый жизненный опыт, и очень общительны — вплоть до заботы о голодных сородичах у вампиров Desmodus rotundus. Но эти преимущества сводятся на нет малыми размерами мозга — дорогая плата за покорение небес. Удивительно, но за десятки миллионов лет ни одна летучая мышь не потеряла способности к полёту и не вернулась к наземному или древесному образу жизни (в фантастической фауне будущего изобретательный ум Д. Диксона нагрезил хищных наземных вампиров, ходящих на передних лапах и хватающих добычу задними, но этот инфернальный образ пока, к счастью, остаётся сугубо гипотетическим и остаётся на совести своего создателя).
Много шуму в своё время наделала так нываемая гипотеза "летающих приматов", согласно которой мегахироптеры Megachiroptera — крылановые — приобрели способность к полёту независимо от прочих летучих мышей — микрохироптеров Microchiroptera, да к тому же возникли из древнейших приматов (Pettigrew, 1986; Pettigrew et al., 1989; Pettigrew et al., 2008). В доказательство приводилось множество доводов, основными из которых был специфический тип нервного соединения сетчатки глаза с верхниками холмиками четверохолмия в среднем мозге — уникальный для приматов, шерстокрылов и крылановых, а также отсутствие эхолокации у подавляющего большинства последних, в отличие от мелких эхолоцирующих рукокрылых. Приводились и другие доказательства независимого возникновения макро- и микрохироптеров.
Читать дальшеИнтервал:
Закладка:
Похожие книги на «Шанс для приматов»
Представляем Вашему вниманию похожие книги на «Шанс для приматов» списком для выбора. Мы отобрали схожую по названию и смыслу литературу в надежде предоставить читателям больше вариантов отыскать новые, интересные, ещё непрочитанные произведения.






![Станислав Дробышевский - Палеонтология антрополога. Книга 1. Докембрий и палеозой [litres]](/books/404766/stanislav-drobyshevskij-paleontologiya-antropologa-thumb.webp)





Обсуждение, отзывы о книге «Шанс для приматов» и просто собственные мнения читателей. Оставьте ваши комментарии, напишите, что Вы думаете о произведении, его смысле или главных героях. Укажите что конкретно понравилось, а что нет, и почему Вы так считаете.